![]()
![]()
![]()
![]()
SPATHES
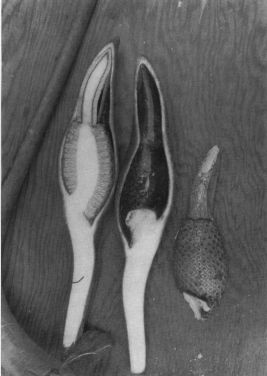
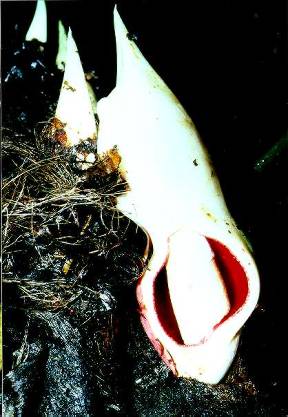
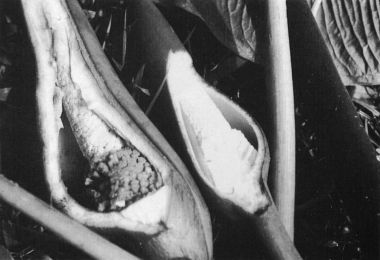
The spathe of P. subg. Philodendron is highly variable in many regards. The spathe of Philodendron is typically coriaceous, and is constructed in such a way that the spathe may open and reclose without major alteration in its shape. The margins are invariably much thinner with the apical portion of the spathe usually closely rolled and frequently acuminate (Fig. 178, P. findens) and the basal portion more conspicuously convolute. Because the margins of the base are more conspicuously overlapped, when the spathe opens the lower portion remains convolute (Fig. 70, P. auriculatum) whereas the apical portion separates to very near the tip. Some species have spathes that open more fully so that the opening is almost rounded (Fig. 298, P. panamense) or broadly elliptic (Fig. 288, P. mexicanum). In such cases more of the inner tube surface and the pistillate spadix is exposed. On the other hand, some species have spathes which open only slightly so that the opening is merely elliptic with the pistillate spadix not visible without peering down through the top of the opening (Fig. 63, P. antonioanum).
Species vary greatly in the extent to which the spathe remains convolute at the base during anthesis. Some species, e.g., P. angustilobum (Fig. 51) and P. mexicanum (Fig. 288), unfold to very near the base. Other species, e.g., P. antonioanum (Fig. 63), do not open very broadly. The spathe tube of P. auriculatum (Fig. 70), P. panamense (Fig. 298), and P. wendlandii (Fig. 451) remains convolute only in the lower half. The spathe of P. davidsonii (Figs. 134, 136) may be convolute only in the lower 1/3 to the lower 2/3 in P. squamicaule. That of P. hebetatum (Fig. 226) is convolute to the upper 1/4 and the spathe tube of P. aromaticum (Fig. 68), P. brenesii (Fig. 89), P. copense (Fig. 110), P. scalarinerve (Fig. 140), P. findens (Fig. 177), P. rothschuhianum (Fig. 340), P. straminicaule (Fig. 360) and P. schottianum (Fig. 371) remain essentially closed to the apex of the tube during anthesis.
Working principally with South American species, Mayo (1986) presented an as yet not fully tested classification of inflorescence types. Eleven inflorescence types were characterized and a key was prepared to separate them. The principal characters were the presence or absence of resin ducts in the inner surface of the spathe. Those with resinous spathes were divided into species with solitary versus species with two or more inflorescences per axil. Those with solitary inflorescences were divided into species with the sterile staminate portion of the spadix equal to or longer than the staminate portion, and visa versa, while those with several inflorescences were divided into species with and without colorful spathe tubes, each group in turn divided into species with constricted and non-constricted spathes.
The non-resinous-spathed species in turn were divided into those species which did or did not have resinous spadices with the resinous-spathed species being further subdivided into species with resin ducts in the staminate portion versus species with resin ducts in either the fertile staminate portion or both sterile and fertile staminate portions. The latter were again subdivided into species with non-functional resin canals versus species with no resin canals and finally the latter group is further subdivided into species with spadices possessing or lacking a terminal sterile zone.
While I believe that this classification may not be completely natural, it demonstrates many evolutionarily important morphological features and shows the complexity of the Philodendron spathe.
Species of P. subg. Philodendron have the spathe tube relatively well demarcated from the blade either by being constricted at the apex or by their contrasting colors, i.e., the spathe tube and spathe blade are differently colored. Some species, e.g. P. crassispathum (Fig. 124), lack any constriction above the tube. Thus the spathe is more or less elliptic. Other species which have a weakly constricted spathe include P. advena (Fig. 40), P. alticola (Fig. 45), P. breedlovei (Fig. 85), P. purulhaense (Fig. 323), P. schottianum (Fig. 368) and P. antonioanum (Fig. 63). Despite these exceptions most members of P. subg. Philodendron have spathes to some extent constricted. The subgenera Meconostigma and Pteromischum on the other hand are not very likely to be constricted or only weakly constricted (Mayo, loc. cit.).
Spathe length is relatively variable depending on the age of the plant and its stage of development. Elongation of the spathe is typically more or less arrested after anthesis but sometimes a considerable amount of elongation takes place in both the peduncle and spathe after anthesis, presumably to accommodate the considerable expansion of the developing fruits. Spathe length varies from as little as 4.5 cm in P. clewelii and 4.8 cm in P. heleniae to 29 cm in P. pterotum and 30 cm in P. warszewiczii. The average maximum length for all species is 16.5 cm. Relatively few species ever exceed 25 cm in length even when in fruit. Additional species that exceed 25 cm include P. davidsonii and P. radiatum var. radiatum. Several additional species have maximum spathe lengths greater than 20 cm. They include the following: P. aromaticum, P. brunneicaule, P. dolichophyllum, P. dressleri, P. ferrugineum, P. findens, P. fortunense, P. lentii, P. llanoense, P. pseudoauriculatum, P. sagittifolium, P. hederaceum ssp. kirkbridei, P. sousae, P. thalassicum, P. tripartitum, and P. verrucosum.
More common are those species which have a maximum spathe length between 15 and 20 cm. Species in this category include: P. advena, P. alticola, P. angustilobum, P. anisotomum, P. annulatum, P. brenesii, P. copense, P. correae, P. cotonense, P. dodsonii, P. fragrantissimum, P. gigas, P. hebetatum, P. hederaceum ssp. hederaceum, P. immixtum, P. jacquinii, P. ligulatum, P. malesevichiae, P. mexicanum, P. microstictum, P. panamense, P. pirrense, P. platypetiolatum, P. purpureoviride, P. purulhaense, P. radiatum var. pseudoradiatum, P. rothschuhianum, P. scalarinerve, P. schottianum, P. smithii, P. squamicaule, P. squamipetiolatum, P. straminicaule, P. strictum, P. tysonii, P. verapazense, P. wendlandii, and P. wilburii var. longipedunculatum.
Almost an equal number have spathes whose maximum lengths are between 10 and 15 cm long. These include: P. albisuccus, P. auriculatum, P. bakeri, P. basii, P. breedlovei, P. brevispathum, P. squamipetiolatum, P. chiriquense, P. ligulatum var. ovatum, P. cotobrusense, P. crassispathum, P. cretosum, P. dominicalense, P. edenudatum, P. folsomii, P. glanduliferum, P. grandipes, P. hammelii, P. heleniae, P. jefense, P. jodavisianum, P. lazorii, P. ligulatum var. heraclioanum, P. morii, P. roseospathum var. roseospathum, P. hederaceum ssp. oxycardium, P. sulcicaule, P. tenue, P. utleyanum, and P. wilburii var. wilburii.
Only a few species have spathes less than 10 cm long. These include P. brewsteriense, P. chirripoense, P. clewelii, P. dwyeri, P. knappiae, P. roseospathum var. angustilaminatum, and P. ubigantupense.
Taxonomically significant but seldom mentioned is the shape of the spathe and the relative disposition of the spadix at anthesis. This is owing to the fact that many species are still poorly known at anthesis, no doubt due to the fact that anthesis in Philodendron lasts only for one or two days for each inflorescence.
One of the features which determine the shape of the spathe at anthesis is the degree to which the spathe opens. Some species, such as P. aromaticum, P. copense, P. dodsonii, P. findens, P. fragrantissimum, P. grandipes, P. hebetatum, P. jodavisianum, P. straminicaule, and P. strictum open only on the blade portion (see discussion above), leaving an oblong-elliptic opening on the blade portion of the spathe only (Fig. 110, P. copense; Fig. 140, P. dodsonii; Fig. 378, P. smithii). In these examples the spadix is either included within the spathe or it is barely exserted, stiffly erect yet held slightly in front of the top edge of the spathe. On the other hand some species, such as P. angustispadix, P. mexicanum, and P. tripartitum may open so broadly as to expose much of the pistillate spadix (Fig. 423 P. tripartitum). It is usually for the most part obscured by the prominently convolute tubular portion of the spadix, as in P. antonioanum, P. aromaticum, P. copense, and P. dodsonii. Those species which have spathes which open more broadly also may have the spadix protruding forward somewhat out of the spathe. See section on Spadix.
The definition of the spathe tube is somewhat inexact but it may be defined as the lower portion of the spathe, that portion of which covers the pistillate portion of the spadix. In reality it usually extends somewhat above the pistillate portion to include the lowermost or sterile portion of the staminate spadix. In cases where the spathe has an obvious constriction it is that portion below the constriction (the balance being the spathe blade) that is referred to as the spathe tube. Though the ending of the spathe tube is imprecise both in terms of its morphology and color, it generally terminates where the greatest constriction of the staminate spadix occurs. This narrowest portion of the staminate spadix usually occurs immediately above the sterile staminate spadix. Functionally, the constriction of the spathe, coinciding with the narrowest portion of the staminate spadix, disallows much of the pollen from the fertile upper portion of the spadix from falling directly into the chamber of the pistillate portion of the spathe. This probably serves more to prevent wastage of the pollen than it does to prevent self-pollination since most species are so markedly protogynous that no self pollination is possible, i.e., the pistils are no longer receptive when the pollen is shed. Not surprisingly those species, e.g., P. advena, P. crassispathum (Fig. 124) and P. purulhaense, which have spathes which are scarcely constricted above the tube also have spadices which are not markedly narrowed above the sterile staminate flowers. It is not known if the pollination behavior of these species differ.
For Philodendron species with a prominent constriction the partially closed spathe during staminal anthesis, coupled with the swollen portion of the staminate spadix immediately below it, probably provide a "pollen well" through which the beetles must squeeze on their departure. This probably ensures a greater coverage of pollen for the departing beetle pollinators.
Spathe tube shape is not particularly interesting. Tubes range in shape from virtually oblong, e.g., in P. heleniae, P. ligulatum, and P. tripartitum, to oblong-elliptic, e.g., in P. aromaticum, P. copense, P. dodsonii, P. findens, P. gigas, P. hebetatum, and P. pseudoauriculatum to elliptic, e.g., P. rothschuhianum and P. smithii and range from only 2 cm long in P. clewelii and to 2.5 cm long in P. grandipes to 13 cm long in P. schottianum and 14 cm long in P. davidsonii and P. warszewiczii. Only a few additional species have spathe tubes longer than 10 cm. These include: P. auriculatum, P. ferrugineum, P. sousae, and P. tysonii. Species with spathe tubes longer than 8 cm include P. advena, P. aromaticum, P. brunneicaule, P. copense, P. dodsonii, P. findens, P. grayumii, P. lentii, P. llanoense, P. malesevichiae, P. panamense, P. straminicaule, and P. verapazense. Even more, 25 species, have spathe tubes greater that 6 cm long. Seventeen species have spathe tubes less than 5 cm long. Even though the spathe tube often remains totally closed at anthesis it is none the less still somewhat expanded owing to the separation caused by the flaring of the spathe blade. Generally this provides considerable room around the spadix to accomodate the pollinating beetles (Figs. 100, 128, 242, 268). At anthesis the spathe tube is considerably larger than before anthesis.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}